Репарация ДНК — это комплекс генетически предопределенных механизмов восстановления повреждений ДНК, возникающих в процессах репликации и рекомбинации ДНК, а также под действием различных факторов, среди которых основными являются следующие:
- Ультрафиолетовое облучение
- Жесткое радиационное облучение
- Депуринизация или апуринизация — спонтанное отщепление азотистых оснований от дезоксирибозных остатков цепи ДНК
- Спонтанное дезаминирование азотистых оснований
Генетическая основа репарационных механизмов заключена в двухцепочечном строении ДНК, которая состоит из двух комплементарных копий. Это обстоятельство позволяет восстанавливать поврежденную полинуклеотидную цепь ДНК благодаря информации заключенной в ее неповрежденном комплементарном аналоге.
Одним из наиболее распространенных дефектов строения ДНК, возникающий при ее репликации является нарушение комплементарности в определенной паре нуклеотидных остатков. Вероятность подобной ошибки при репликации ДНК составляет около 1:106 нуклеотидных пар. Однако, вовлеченная в процесс репликации, специфическая ДНК-полимераза дельтаспособна вырезать последний нуклеотид, растущей дочерней цепи ДНК в том случае когда он не комплементарен соответствующему нуклеотиду матричной цепи. Этот процесс коррекции ошибки, допущенной ДНК-полимеразой бета, приводит к понижению вероятности нарушений комплементарности до 1:107 нуклеотидных пар. Однако, в некоторых случаях этот механизм не срабатывает и дальнейшая коррекция нарушений комлементарности цепи дочерней ДНК рализуется механизмами системы репарации, которые в конечнем итоге снижают вероятность нарушений комлементарности до 1:109 и более нуклеотидных пар.
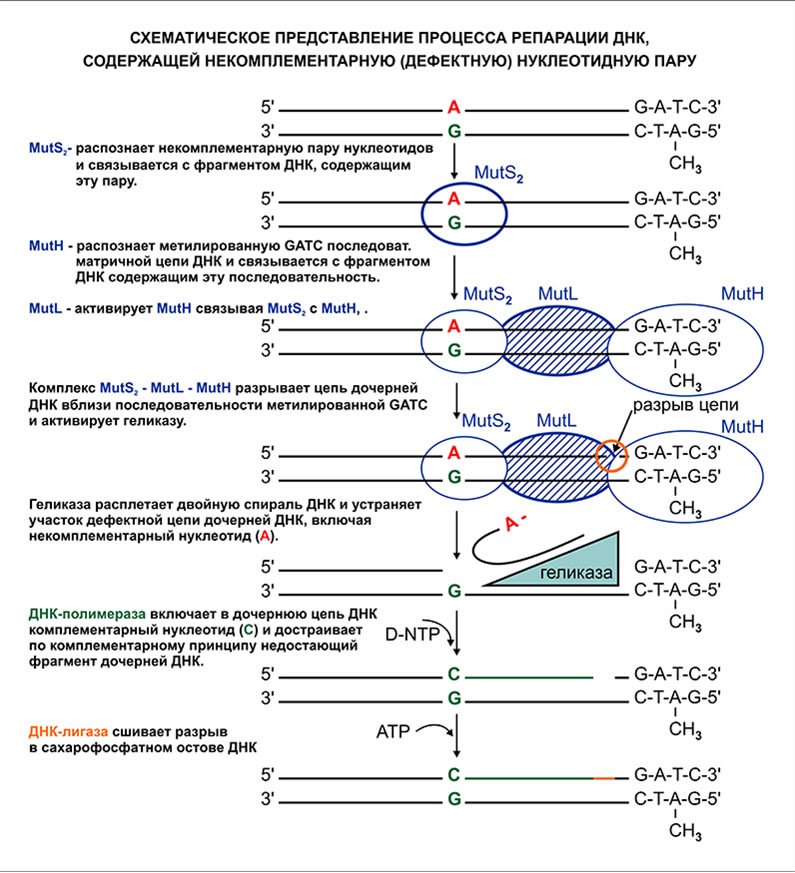
Репарация некомплементарных нуклеотидных пар происходит исключительно на дочерней цепи ДНК. Ферменты участвующие в процессе репарации распознают матричную цепь ДНК по метилированным остаткам адениновых оснований в тетрануклеотидных последовательностях -G-A(СН3)-T-C-. В связи с этим, система репарации должна выявить и исправить ошибку репликации до метилирования адениновых оснований дочерней цепи ДНК.
Репарация ДНК с нарушениями комплементарности нуклеотидных пар, возникающих в процессе репликации ДНК включает в себе следующие объязательные этапы:
- Выявление фрагмента двухспиральной ДНК, содержащей некомплементарную (дефектную) пару нуклеотидов.
- Устранение дефектного фрагмента дочерней цепи ДНК, включая некомплементарный нуклеотид.
- Включение в дочернюю цепь ДНК комплементарного нуклеотида и восстановление ранее устраненного фрагмента ДНК.
- Восстановление непрерывности полимерной цепи посредством замыкания сахарофосфатного остова дочерней цепи ДНК.
Ферменты задействованные в репарации ДНК
MutS2 — фермент, который распознает участок ДНК, содержащий дефекную пару нуклеотидов. Представляет из себя гетерологический димер. Связывается с участком ДНК, включающим в себе некомплеиментарную пару нуклеотидов и при этом, захватывает около 20 нуклеотидных пар, деформируя двойную спираль ДНК.
MutH — фермент, который распознает и связывается с ближайшим к дефектной паре нуклеотидов участоком цепи матричной ДНК, содержащей метилированную по адениновому основанию тетрануклеотидную последовательность GATC, которая может находится на растоянии 1000 и более нуклеотидов от дефектной пары.
MutL — белок, который связывает между собой MutS2 и MutH, образуя активный ферментативный комплекс MutS2 — MutL —MutH, способный разрезать цепь дочерней ДНК вблизи метилированной последовательности GATC.
Геликаза — фермент, активируемый комплексом MutS2 — MutL — MutH. Он расплетает двухспиральную ДНК, начиная с точки разрыва в области локализации последовательности GATCпосле чего, отрезает и удаляет фрагмент дочерней цепи ДНК, захватывая при этом дефектный некомплементарный нуклеотид.
ДНК-полимераза — фермент, включающий в цепь дочерней ДНК вместо удаленного геликазой дефектного нуклеотида, комплементарный нуклеотид, после чего, достраивает эту цепь по принципу комплементарности до точки разрыва вблизи метилированной GATC— последовательности матричной цепи ДНК.
ДНК-лигаза — фермент, который сшивает разрыв сахарофосфатного остова дочерней цепи ДНК вблизи метилированной GATCпоследовательности матричной ДНК.
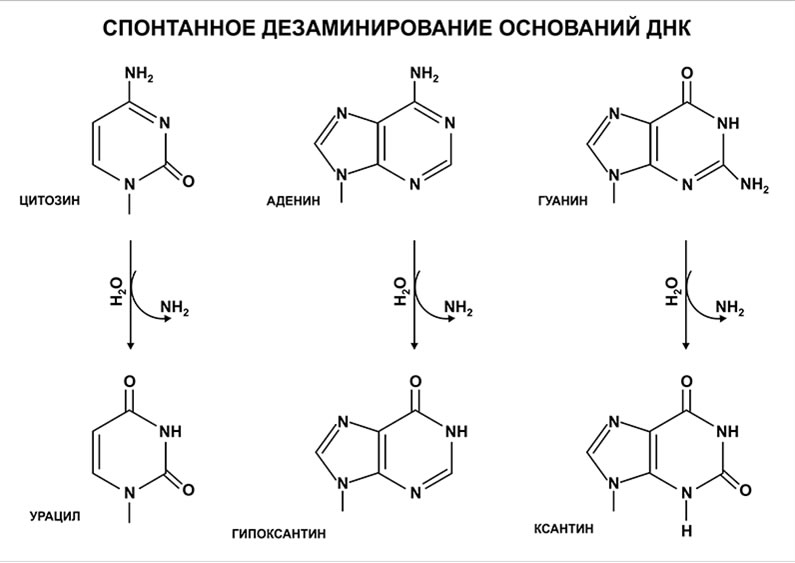
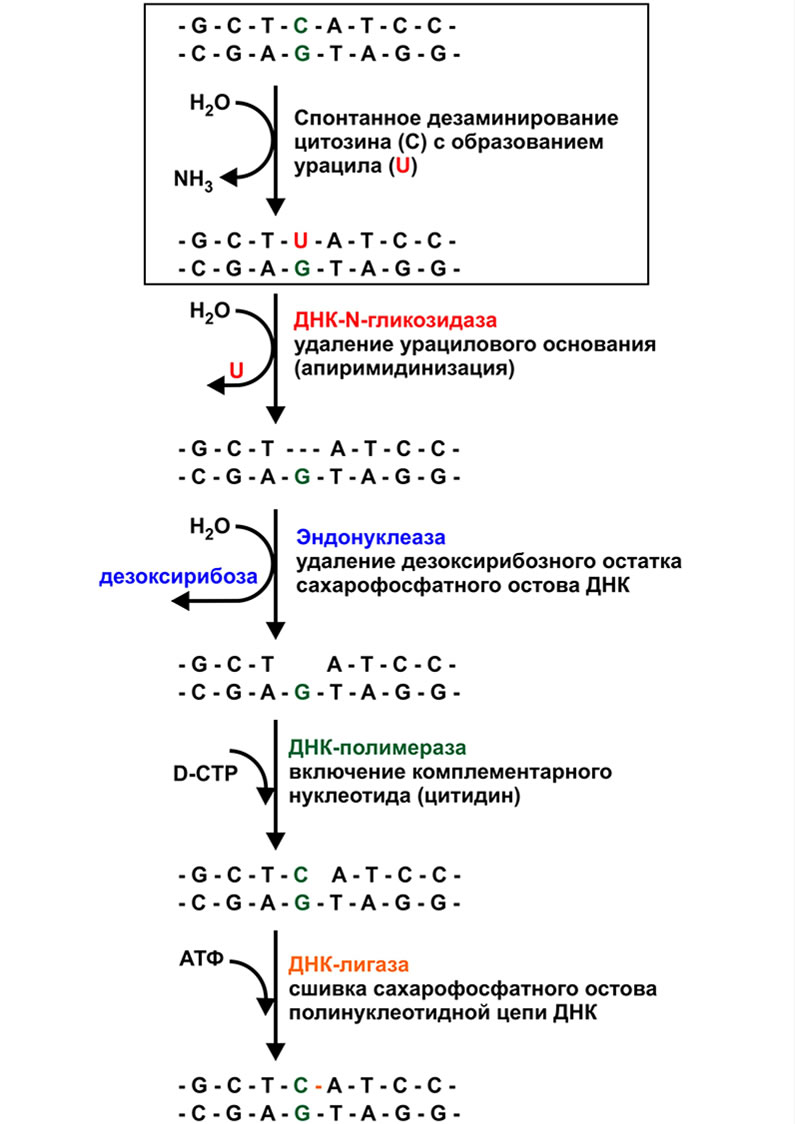
Одним из редко встречаемых типов повреждений ДНК является спонтанное дезаминирование азотистых оснований согласно представленному рисунку.
Все продукты дезаминирования азотистых оснований ДНК — урацил, гипоксантин и ксантин не типичны для структуры ДНК и легко распознаются ферментами системы репарации.
В восстановлении данного типа повреждения ДНК задействованы следующие ферменты:
- ДНК-N-гликозидаза, которая удаляет дезаминированное основание /осуществляет апиримидинизацию (AP) нуклеотида.
- AP-эндонуклеаза, которая удаляет дезоксирибозный остаток
- ДНК-полимераза бета, которая включает в структуру ДНК комплементарный нуклеотид.
ДНК-лигаза, которая сшивает разрыв сахарофосфатного остова ДНК.